DNA can loop to bind tetramers of repressor at 01-02 or 03-01 or both loops; each loop
Ý repression 30-fold
looping can provide repression of 70-fold compared to that in single binding of 17-fold
Pap Operon
ability to turn pili formation on/off since pili help adhesion to walls but also lead to phagocytosis
signals – increased temperature or lousy nutrition
Þ on
regulation – protein binding to specific sites on DNA, looping, methylation of GATC sites on the DNA proper
Attenuation
relies on simultaneous transcription and translation
involves speed of ribosome movement down the mRNA molecule and alternative base pairing
proportional to levels of tRNA for the amino acid synthesized by the transcribed genes
trp similar to his (see figure below):
attenuator stops transcription
attenuator can be modified by changes in the secondary structure of the mRNA
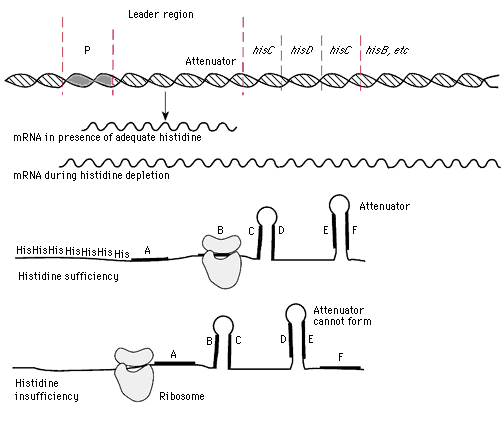
Schematic representation of the control of transcription by the process of attenuation. The example chosen is the his operon of Escherichia coli. How attenuation works is fascinating. The leader region is always transcribed and translated into a small oligopeptide. The peptide near the attenuator site has a string of seven his codons. Movement of the first ribosome coming behind the polymerase is drastically affected by the supply of charged his tRNA. If there is an adequate supply, the ribosome is not delayed, and an attenuator loop forms in the mRNA, causingranscription to terminate. With a shortage of histidine, the first ribosome gets hung up over the his codons, and the attenuator loop is not formed, because alternate loops form. As a result, transcription proceeds, the complete his mRNA is made, and the biosynthetic enzymes can be made in large quantities. the upper portion of the figure illustrates the difference in transcription of the his operon in histidine sufficiency and insufficiency; the lower two diagrams depict the molecular mechanism of attenuation.